新疆主栽海岛棉品种(系)遗传多样性的SSR分析 
2. 新疆溢达纺织有限公司, 乌鲁木齐, 830000
3. 黄冈师范学院化学与生命科学学院, 黄冈, 438000
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2013 年, 第 11 卷, 第 18 篇 doi: 10.5376/mpb.cn.2013.11.0018
收稿日期: 2013年03月20日 接受日期: 2013年05月08日 发表日期: 2013年06月18日
引用格式(中文):
郭欢乐等, 2013, 新疆主栽海岛棉品种(系)遗传多样性的SSR分析, 分子植物育种(online), 11(18): 1123-1130 (doi: 10.5376/mpb.cn.2013.11.0018)
引用格式(英文):
Guo et al., 2013, Genetic Diversity Analysis of Widely Cultivated Sea-island Cultivars (Lines) in Xinjiang Using SSR Markers, Fenzi Zhiwu Yuzhong (online) (Molecular Plant Breeding), 11(18): 1123-1130 (doi: 10.5376/mpb.cn.2013.11.0018)
为了揭示新疆主栽海岛棉品种(系)的遗传差异,更好地指导新疆海岛棉育种,利用187对覆盖棉花全基因组的SSR引物对20个新疆海岛棉主栽品种(系)进行了遗传多样性分析。共有55对引物表现多态性,占总引物数的29.4%,共得到多态性位点97个,平均每对引物产生1.8个多态性位点。等位基因变异的多态性信息含量(PIC)在0.095~0.829之间,平均为0.560。成对品种间遗传相似系数在0.18~0.93之间,平均成对相似系数为0.44。聚类结果显示阈值为0.38时,20个品种(系)可分为两大类;在阈值为0.43和0.48时,两大类材料可分别分为两个亚类。本研究结果表明新疆主栽海岛棉品种(系)的遗传基础较宽,生产风险较小。
棉花是重要的经济作物,也是纺织业重要的原料来源。在棉花纤维产量中,虽然长绒棉(海岛棉)产量占棉花总产量不到10%,但是由于长绒棉具有的纤维长度长、丝光好和强力高等优点,是高档面料的重要来源,在纺织业中占有不可替代的作用。随着近年来人民生活水平的提高,高档面料的需求日趋旺盛,同时随着纺织业的快速发展以及国际纺织市场的激烈竞争,对棉纤维特别是长绒棉的品质和产量都提出了新要求。目前中国长绒棉年需求量在2.0×108 kg左右,而长绒棉年产量多数年份都在1.5×108 kg以下,缺口较大,并且棉纤维的品质已经越来越难以满足纺织工艺越来越高的要求,特别是在120支高档纱方面表现更为明显(丁胜等, 2009, 中国种业, (12): 17-19)。由于长绒棉生产具有生产周期长、市场风险大和经济效益低等方面的不利因素,严重影响了棉农的种植积极性(曹吉强等, 2011, 中国棉花, 38(5): 6-8),导致近年来长绒棉的种植面积波动较大,进一步加剧了供求矛盾,这些状况使中国棉花产业面临日益严峻的局面。因此,积极探讨海岛棉品种遗传背景,培育高产优质新品种具有重要意义。
遗传多样性是指某一物种种内的不同种群之间或者一个种群内部的不同个体之间的遗传变异,遗传多样性有助于了解物种群体间或者群体内的变异水平,遗传结构等信息,是农作物品种遗传改良的基础。遗传多样性研究有助于了解品种的亲缘关系和群体结构等信息,对作物品种资源的保护、利用及指导其遗传育种具有重要意义。
分子标记技术由于具有不受环境因素的影响,结果稳定可靠等优点, 近年来被广泛应用于遗传多样性分析。国内利用分子标记技术对棉花进行遗传多样性分析,近年来也取得了较快进展。徐秋华等利用RAPD技术分析了长江和黄河流域两棉区的91个陆地棉品种,表明两棉区品种的遗传多样性水平相当(徐秋华等, 2001)。姜伟等利用ISSR技术将48份棉花种质划分为4大类,认为材料间的遗传多样性较为丰富(姜伟等, 2008)。李金荣等利用SSR技术对14个海岛棉材料进行聚类分析,将该实验材料聚为4类(李金荣等, 2009)。李武等和Wang等分别利用SRAP技术和SSR技术对我国引进海岛棉以来培育的36个品种以及20个国外品种进行了遗传多样性分析,结果显示聚类结果与系谱具有一定的相符性,不同的标记技术分析结果也有一定的差异(李武等, 2008; Wang et al., 2011)。吴大鹏等利用SSR技术对来自四个国家的20份海岛棉材料进行分析,将该材料分为4类(吴大鹏等, 2011)。武路云等利用SRAP技术对168份国内外海岛棉材料进行遗传多样性分析的结果表明,海岛棉材料差异大,遗传多样性丰富(武路云等, 2012)。目前,在作物遗传多样性研究中,应用较多的分子标记技术类型有RFLP、RAPD、AFLP和SSR等。其中SSR标记具有多态性高、共显性分离、位点专化性以及标记覆盖整个基因组且分布均匀和DNA样本用量少等优点(朱四元等, 2006), 被认为是研究群体遗传标记变异的最好标记之一。
目前国内对海岛棉的遗传多样性研究多是将国内外种质资源进行比较分析,缺少对新疆主栽海岛棉品种(系)的遗传多样性分析报道。本研究利用SSR标记通过对目前新疆主栽的海岛棉品种(系)共20个材料进行遗传多样性检测,旨在从分子水平上对品种遗传变异的大小和遗传结构进行分析,从而揭示该海岛棉材料的遗传差异及其相互关系,进而为科学地评价其遗传潜力、保护和利用种质资源提供一定的理论依据。
1结果与分析
1.1 SSR引物多态性分析
在187对引物中共有55对引物有多态性,占总引物数的29.4%,共得到多态性位点97个,平均每对引物产生1.8个多态性位点,其中引物BNL1026扩增出多态性条带最多(4条),表明该引物所在位点变异较为丰富,能较好地反映出材料的遗传多样性。等位基因变异的多态性信息含量(PIC)在0.095~0.829之间,平均为0.560。本实验得到有多态性的55对引物中,共有26对已经定位到棉花染色体上,除了5号、9号、13号、17号、24号染色体外其余染色体均有分布,分布情况比较均匀(表1)。在15号和19号染色体上分布有多态性引物3个,说明该染色体上存在较多的变异位点。
 表1 多态性引物的染色体定位及其PIC值和重复序列 Table 1 Chromosome localization, PIC value and motif of polymorphic primers |
在55对有多态性的引物中,有41对已经获取了其重复序列(http://www.cottonmarker.org/)。从表1可以看出重复序列的长度与PIC值并无直接关联,重复序列的长度并未随着PIC值的增大而增长或缩短,而是长度随机分布,在各个PIC值区段均有较长和较短序列存在。在各种类型的基序中,二核苷酸和三核苷酸出现次数最多,分别为13次和15次,五核苷酸出现次数较少,只有1次,长度大于7的出现4次(图1),说明二核苷酸和三核苷酸与多态性的产生有较大关联。
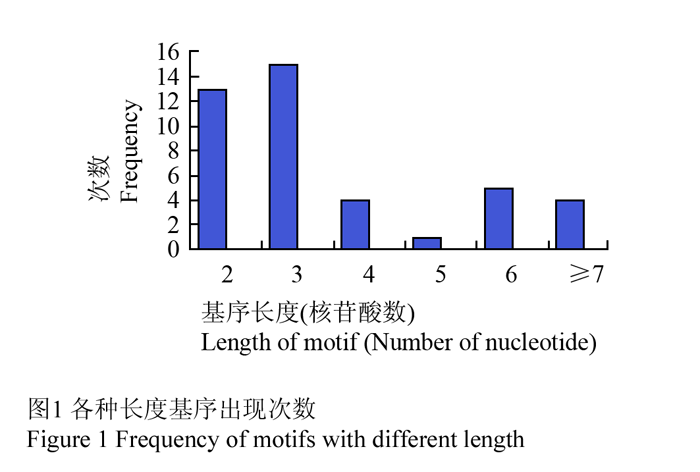 图1 各种长度基序出现次数 Figure 1 Frequency of motifs with different length |
1.2基于SSR的遗传多样性分析
1.2.1成对相似系数
利用软件NTSYSpc2.1将SSR分子标记在20个品种(系)中的多态性数据计算出两两材料间的相似系数。由表2可知,试验材料间遗传相似系数在0.18~0.93之间,平均为0.44,其中71%的材料间相似系数小于0.5,表明材料间遗传差异较大。其中5917和新海27号之间遗传相似系数最小(0.18),表明两者亲缘关系较远。新海35号和新海29号的遗传相似系数最大(0.93),表明两者亲缘关系很近,遗传背景非常相似。
 表2 材料间成对遗传相似系数 Table 2 Pairwise genetic similarity coefficients of these materials |
1.2.2 SSR标记聚类分析
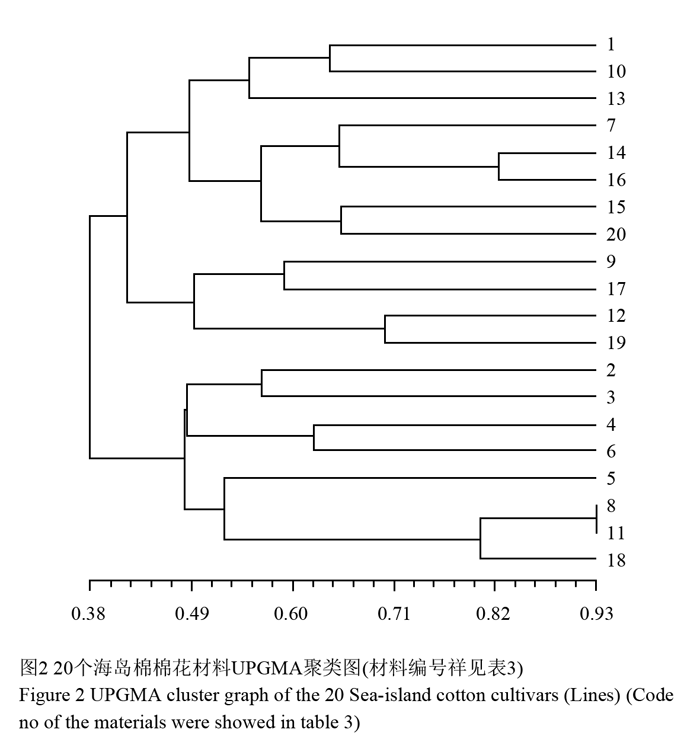
聚类分析采用类平均法(UPGMA)进行。由图2可知在阈值约为0.38时可将该材料分为两个大类;第一类包括12个材料:新海15号、新海33号、塔07-152、新海28号、DJ-1、K388、阿长-599、S0717、新海31号、DJ-07-136、H3549和5917。这12个材料在阈值为0.43时又可分为两个亚类,第一亚类包括新海15号、新海33号、塔07-152、新海28号、DJ-1、K388、阿长-599和S0717。第二亚类包括H3549、5917、新海31号和DJ-07-136。第二大类包括8个材料:新海21号、新海24号、新海25号、新海27号、新海26号、新海29号、新海35号和天长2号。第二大类在阈值约为0.48时又可分为两个亚类,第一亚类包括新海21号、新海24号、新海25号和新海27号。第二亚类包括新海26号、新海29号、新海35号和天长2号。由图2可知第一大类和第二大类之间遗传相似系数较小,表明其遗传背景差异较大。
2讨论
2.1聚类结果与系谱分析
本实验所用材料除新海31号和天长2号外,其余均得到其系谱来源(黄滋康, 2007, 中国农业出版社, pp.341-342; 洪梅等, 2008a, 中国棉花, 35(6): 26; 李尔文等, 2001, 中国棉花, 28(2): 28; 宁新民等, 2005, 中国棉花, 32(11): 26; 王献礼等, 2005, 中国种业, (9): 54; 洪梅等, 2008b, 中国棉花, 35(8): 21; 余力等, 2010, 中国棉花, 37(10): 30; 李尔文等, 2004, 中国棉花, 31(1): 31; 江清等, 2001, 中国棉花, 28(4): 30; 李尔文等, 2002, 中国棉花, 29(6): 22)。本实验材料间聚类结果与前人研究结果基本相符但有一定差异,这可能是由于选用的引物不同,特异性扩增位点不同等原因造成的。从聚类结果和系谱分析结果对比来看,两者基本相符并有一定差异,这与前人的研究结果一致。从系谱来源上看,第一大类材料总体上比第二类含有更多的埃及型海岛棉成份,因此在分子水平上可以将其区分开。在第一大类中,新海15号、新海33号和塔07-152聚在一起,这与亲本来源相符,其亲本都含有新海10号和埃及棉A杂交铃,其中新海33和塔07-152的亲本之一新海8号与三者的共有亲本新海10号来源相同,因此这3个材料可以聚在一起。DJ-1由新海28号优选得出,因此它们聚在一起。阿长599和S0717有共同的亲本新海21号,阿长-599的另一亲本新海18号与S0717的另一亲本新海14号具有相似来源,因此阿长-599和S0717可以聚在一起。在第二大类中,新海26号和新海35号具有相同的母本来源,亲缘关系较近,因此它们聚在一起。实验结果也有与系谱分析不一致的情况,如新海21号是塔07-152、S0717、阿长-599的亲本来源,但是新海21号并没有在聚类结果中和它们聚在同一大类中,这可能是其后代产生了较严重的分离,新海21号的遗传物质被保留的较少。DJ-07-136是由新海21号系统选育出的品系,也没有和新海21号聚在同一大类中,可能在选育过程中发生了较多自然变异或基因重组,这也可以从另一方面揭示DJ-07-136可以脱离新海21号成为一个新的品系的原因。新海21号和新海33号出现在两个大类中,新海33号由新海17号辐射后选育而来,而新海21号和新海17号有相同的亲本来源,只是亲本组合方式不同((新海8号×吉扎75)F2×(新海10号×A杂交铃)F2→新海21;(新海8号×吉扎75)F1×(新海10号×A杂交铃)F1→新海17号),这可能是在选育过程中发生了严重分离并且发生较多突变再加上人为选择引起的。聚类结果和系谱分析的差异也可以从另一方面说明杂交选育可以有效改变材料的遗传背景。
2.2分子标记分析与系谱分析
系谱分析经常用来进行遗传多样性分析,从材料来源上分析其亲缘关系的远近。从前人和本实验研究结果来看,系谱分析并不能与DNA聚类分析完全吻合,往往有一定的差异。在育种过程中,育种者经常采取各种方法来开展工作,如采取两个亲本杂交,多亲本杂交以及回交等等,培育出的品种经常有多个亲本来源。这些亲本对子代遗传物质的贡献往往差别较大,典型的如回交育种,选育的子代拥有受体亲本的绝大部分基因,而供体亲本只对选育的品种贡献个别有利基因。因此虽然供体亲本是两个亲本之一,但是在DNA聚类分析上并不能一定将供体亲本和子代聚在一起。结合分子标记技术的聚类分析能够从DNA水平上分析品种间的差异,其聚类结果能更加真实地反映品种间的遗传差异,因而分析结果具有更强的说服力。系谱分析偏向于说明材料来源,DNA聚类分析则偏向于揭示材料DNA序列上的差异,因此将二者结合起来进行分析,才能对材料的遗传多样性有更加全面的了解。
2.3遗传多样性与棉花育种
刘文欣等对建国以来166个具有代表性的棉花主栽品种进行遗传多样性研究,结果显示在我国主栽棉花品种中,海岛棉品种遗传基础比陆地棉品种狭窄,我国自育品种遗传基础比国外引进品种狭窄,陆地棉杂交种的遗传基础比常规品种窄,上世纪80年代以后育成的陆地棉品种遗传基础比70年代的品种狭窄(刘文欣等, 2003)。赵战胜等利用SSR标记对北疆早熟陆地棉品种进行遗传多样性分析结果表明,该地区陆地棉遗传多样性狭窄(赵战胜等, 2012) 李武等和陈光等研究均表明,我国海岛棉品种总体上来看遗传多样性丰富,但新育成的品种遗传多样性仍然出现变窄的趋势(李武等, 2008; 陈光等, 2005)。王献礼等和何良荣等分别利用系谱分析的研究结果表明新疆海岛棉的遗传基础在逐渐变窄(王献礼等, 2002, 中国种业, (12): 21-22; 何良荣等, 2002, 中国棉花, 29(6): 8-9)。
总结前人研究结果可知,我国现有棉花品种遗传基础正在趋于狭窄,特别是陆地棉已经出现明显的育种瓶颈,这种现象已经严重制约了棉花育种工作的进展。近年来我国棉花产量并没有较大突破,这与棉花品种遗传背景的相似性不无关联。
在育种材料的选择上,育种者往往选择那些性状优良的材料作为亲本,结合各种育种方法进行选择,性状较差的材料则不断被淘汰,从而形成了一些骨干育种亲本,导致育成的品种往往具有相似的遗传背景。这一现象从本研究中也可以发现,如军海1号是新海8号和新海10号的亲本之一;新海10号是新海14号、新海15号、新海17号、新海21号、新海24号和新海29号等材料的直接或者间接来源;新海8号是新海18号、新海25号、新海21号、新海17号、新海33号等材料的直接或者间接来源;新海21号是塔07-152、S0717、DJ-07-136和阿长-599等材料的直接或者间接来源。从来源相似的材料中所选育的品种,由于亲本遗传基础狭窄,必然难以产生有突破性的新品种。品种遗传背景单一也可以使某些病菌得到选择,产生侵染面较广的病菌生理小种,这对我国棉花生产也是一个巨大的潜在威胁。因此,积极收集和利用新的育种材料,采取各种方法如:引进国外品种资源;栽培棉与野生棉杂交;海岛棉与陆地棉之间杂交;人工培育突变材料等拓宽现有品种遗传背景极具现实意义。
在杂交种选育过程中,亲本的选择十分重要。经验证明亲缘关系远的亲本较有可能创造优良的组合,但是亲缘关系太远的亲本产生的后代则容易发生不育现象。通过遗传多样性分析可以将来源不同的材料归类,从而确定材料间亲缘关系的远近,这就为亲本的选择指明了方向,可以有效避免育种工作的盲目性。在玉米育种工作中,已将玉米各品系分为五类,在这五类之间进行相互组配来选育杂交种,从而提高选育效率(王懿波等, 1998)。近年来我国棉花产量没有太大进展,而优良杂交种的选育是一条能快速提高产量的途径。笔者认为,棉花杂交种的选育也可以参照上述方法,将现有棉花育种资源整理归类,在各个种类间进行杂交组配来提高选育效率。本研究利用覆盖棉花全基因组的187对SSR引物对20个海岛棉材料进行了遗传多样性分析,聚类结果显示在相似系数为0.38时可以将材料分为两类,在今后的品种选育过程中可以在两类之间进行杂交组配。
3材料方法
3.1供试材料
实验所用材料为20份新疆海岛棉主栽品种和一些品系,由新疆溢达纺织有限公司提供(表3)。
 表3 20个供试棉花品种(系) Table 3 20 cotton varieties(lines) used for syudy |
3.2方法
3.2.1棉花基因组DNA的提取
所有参试材料于2011年冬在华中农业大学作物遗传改良国家重点实验室,利用种子培育无菌苗,取刚展开的嫩叶采用CTAB法进行DNA的提取(宋国立等, 1998)。
3.2.2 SSR引物来源,PCR扩增和聚丙烯酰胺凝胶电泳
本实验所用SSR引物共有3个来源:(1)根据已经公布的棉花遗传种间连锁图谱,每10 cM挑取一个标记(Yu et al., 2011);(2)白静等(2012)筛选的杂交棉SSR核心引物;(3)本实验室构建海岛棉种内图谱时筛选的多态性丰富、带型清晰的SSR引物,总数为187对。分别在20个品种(系)间进行SSR-PCR反应。PCR反应总体积为10 μL, PCR反应体系:DNA模版(10 ng/μL) 2.5μL;正向引物(25 μmol/L) 0.16 μL;反向引物(25 μmol/L) 0.16 μL;dNTP (10 mmol/L) 0.25 μL;Taq Buffer (10×) 1.00 μL;MgCl2 (25 mol/L) 0.8 μL;Taq酶(5 U/μL) 0.16 μL;ddH2O 4.97 μL。
扩增仪器型号为Bio-Rad Mycycler,反应程序为95℃预变性5 min,94℃变性50 s,56℃退火45 s,72℃延伸1 min,循环34次,72℃延伸5 min。扩增产物经变性后在DY-12型电泳仪上在6%聚丙烯酰胺变性凝胶中80 W恒功率电泳1~1.5 h,然后银染固定。
3.2.3数据统计及分析
在凝胶上有多态性位点的条带上,有扩增位点的,记为“1”,没有的记为“0”,缺失的记为“2”进而形成原始数据阵。采用NTSYSpc2.1软件进行聚类分析,获得相似系数矩阵和聚类图。多态性信息含量(PIC)计算公式为:
其中Pj为第i位点第j等位变异的频率(Anderson et al., 1993)。
作者贡献
郭欢乐、张明菊是本研究的实验设计和实验研究的执行人;郭欢乐和刘霞完成数据分析,论文初稿的写作;张明菊参与实验设计,试验结果分析;林忠旭是项目的构思者及负责人,指导实验设计,数据分析,论文写作与修改。全体作者均阅读并同意最终的文本。
致谢
本研究由国家自然科学基金项目(30871559)资助。感谢匿名同行评审人的修改建议和评审建议。
参考文献
Anderson J.A., Churchill G.A., Autrique J.E., Tanksley S.D., and Sorrells M.E., 1993, Optimizing parental selection for Genetic linkage maps, Genome, 36(1): 181-186
http://dx.doi.org/10.1139/g93-024
PMid:18469981
Bai J., Nie Y.C., Lin Z.X., Guo X.P., Zhang X.L., Wang B., and Liu C.X., 2012, Screening and evaluation of SSR core primers for indentification of cotton hybrids, Mianhua Xuebao (Cotton Science), 24(3): 207-214 (白静, 聂以春, 林忠旭, 郭小平, 张献龙, 王斌, 刘传祥, 2012, 棉花杂交种SSR核心引物的筛选与评价, 棉花学报, 24(3): 207-214)
Chen G., Du X.M., Lu D.B., Zhou Z.L., and Liu G.Q., 2005, Genetic diversity of sea island cotton using SSR markers, Zhiwu Yichuan Ziyuan Xuebao (Journal of Plant Genetic Resources), 6(2): 135-139 (陈光, 杜雄明, 卢东柏, 周忠丽, 刘国强, 2005, 利用SSR分子标记进行海岛棉遗传多样性研究, 植物遗传资源学报, 6(2): 135-139)
Jiang W., He J.M., Lu J.N., and Zhong X.W., 2008, Analysis of the genetic diversity of some main cultivated cotton populations in China, Nongye Shengwu Jishu Xuebao (Journal of Agricultural Biotechnology), 16(1): 127-133 (姜伟, 何觉民, 陆建农, 钟孝威, 2008, 中国主要棉花栽培群体的遗传多样性分析, 农业生物技术学报, 16(1): 127-133)
Li J.R., Wang X.G., Zhu Y.J., Zhang X.Y., and Zhang W., 2009, Cluster analysis on 14 sea-island cotton by SSRs, Xinjiang Nongye Kexue (Xinjiang Agricultural Sciences), 46(2): 237-241 (李金荣, 王小国, 朱永军, 张西英, 张薇, 2009, 利用SSR标记对14个海岛棉品种的聚类分析, 新疆农业科学, 46(2): 237-241)
Li W., Ni W., Lin Z.X., and Zhang X.L., 2008, Genetic diversity analysis of Sea-Island cotton cultivars using SRAP markers, Zuowu Xuebao (Acta Agronomica Sinica), 34(5): 893-898 (李武, 倪薇, 林忠旭, 张献龙, 2008, 海岛棉遗传多样性的SRAP标记分析, 作物学报, 34(5): 893-898)
Liu W.X., Kong F.L., Guo Z.L., Zhang Q.Y., Peng H.R., Fu X.Q., and Yang F.X., 2003, Ananalysis about genetic basis of cotton cultivars in China since 1949 with molecular markers, Yichuan Xuebao (Acta Genetica Sinica), 30(6): 560-570 (刘文欣, 孔繁玲, 郭志丽, 张群远, 彭惠茹, 付小琼, 杨付新, 2003, 建国以来我国棉花品种遗传基础的分子标记分析, 遗传学报, 30(6): 560-570)
Song G.L., Cui R.X., Wang K.B., Guo L.P., Li S.H., Wang C.Y., and Zhang X.D., 1998, A rapid improved CTAB method for extraction of cotton genomic DNA, Mianhua Xuebao (Cotton Science), 10(5): 273-275 (宋国立, 崔荣霞, 王坤波, 郭立平, 黎绍惠, 王春英, 张香娣, 1998, 改良CTAB法快速提取棉花DNA, 棉花学报, 10(5): 273-275)
Wang X.Q., Feng C.H., Lin Z.X., and Zhang X.L., 2011, Genetic diversity of sea-island cotton (Gossypium barbadense) revealed by mapped SSRs, Genet. Mol. Res., 10(4): 3620-3631
http://dx.doi.org/10.4238/2011.December.8.5
PMid:22183945
Wang Y.B., Wang Z.H., Lu L.X., Wang Y.P., Zhang X., and Tian Z.Y., 1998, Studies on maize germplasm base, division of heterosis groups and utilizing models of heterosis in China, Yumi Kexue (Maize Sciences), 6(1): 9-13 (王懿波, 王振华, 陆利行, 王永普, 张新, 田曾元, 1998, 中国玉米种质基础、杂种优势群划分与杂优模式研究, 玉米科学, 6(1): 9-13)
Wu D.P., Fang X.X., Cui R.G., Chen J.H., and Zhu S.J., 201l, Genetic relationship and diversity of the upland cotton germplasms from different cotton producing countries using SSR markers, Mianhua Xuebao (Cotton Science), 23(4): 291-299 (吴大鹏, 房嫌嫌, 崔闰根, 陈进红, 祝水金, 201l, 国内外陆地棉品种资源的亲缘关系和遗传多态性研究, 棉花学报, 23(4): 291-299)
Wu L.Y., Liu F., Jin M.K., Kong X.J., Wang K.B., and Zhou R.Y., 2012, Genetic diversity analysis of introduced self-bred sea-island cotton (Gossipium barbadense L.) varieties based on SRAP marks, Nanfang Nongye Xuebao (Joural of Southern Agriculture), 43(7): 901-906 (武路云, 刘方, 靳明凯, 孔祥军, 王坤波, 周瑞阳, 2012, 海岛棉部分引进和自选品种遗传多样性的SRAP分析, 南方农业学报, 43(7): 901-906)
Xu Q.H., Zhang X.L., and Nie Y.C., 2001, Genetic diversity evaluation of cultivars (G. hirsumtum L.) from the Changjiang river valley and yellow river valley by RAPD markers, Yichuan Xuebao (Acta Genetica Sinica), 28(7): 683-690 (徐秋华, 张献龙, 聂以春, 2001, 长江、黄河流域两棉区陆地棉品种的遗传多样性比较研究, 遗传学报, 28(7): 683-690)
Yu Y., Yuan D.J., Liang S.G., Li X.M., Wang X.Q., Lin Z.X., and Zhang X.L., 2011, Genome structure of cotton revealed by a genome-wide SSR genetic map constructed from a BC1 population between gossypium hirsutum and G. barbadense, BMC Genomics, 12: 15
http://dx.doi.org/10.1186/1471-2164-12-15
PMid:21214949 PMCid:3031231
Zhao Z.S., Yu S.X., Fan S.L., Zhang W.G., Pang C.Y., and Mei Y.J., 2012, Analysis of genetic diversity of early maturing upland cotton varieties in northern Xinjiang, Mianhua Xuebao (Cotton Science), 24(6): 473-480 (赵战胜, 喻树迅, 范术丽, 张卫国, 庞朝友, 梅拥军, 2012, 北疆早熟陆地棉品种的遗传多样性分析, 棉花学报, 24(6): 473-480)
Zhu S.Y., Chen J.X., Liu A.Y., Li R.L., Yan Y.W., and Tang H.M., 2006, Genetic diversity analysis and identification of insect-resistant cottons based upon SSR markers, Hunan Nongye Daxue Xuebao (Hunan Agricultural University (Natural Sciences)), 32(5): 469-472 (朱四元, 陈金湘, 刘爱玉, 李瑞莲, 严跃文, 唐海明, 2006, 利用SSR标记对不同类型抗虫棉品种的遗传多样性分析, 湖南农业大学学报(自然科学版), 32(5): 469-472)